

Каспийское море, формально являющееся величайшим в мире солоновато-водным озером, по совокупности своих гидрологических и гидрохимических черт не демонстрирует каких-либо специфических отличий от настоящих внутриматериковых морей. Большое практическое значение Каспийского моря в отношении рыболовства и добычи минерального сырья, отчлененность от океана и возможность подсчета полного солевого баланса и изучения судьбы отдельных химических элементов придают особый интерес химическому изучению этого озера-моря. Тем не менее, портал Qazaqstan Tarihy решил ограничиться более узкой задачей, а именно освещением общего гидрохимического облика Каспийского моря, каким он представляется на основании научных изысканий, имевших место в 1933 году.
Научные исследования по химии Каспийского моря, сделанные до 1930-х годов, заключались в основном в отдельных полных химических анализах воды Каспийского моря, определениях хлора, удельного веса, сероводорода, кислорода, отчасти рН. Подобные работы были произведены главным образом А.А. Лебединцевым, Н.М. Книповичем и В.П. Жузе. На их работах, а также (в большей степени) на работах С.В. Бруевича и основывается данная статья.
В августе 1933 году Государственным Океанографическим институтом были произведены гидрохимические работы на двух широтных размерах в Южном Каспии, давшие возможность построения схемы вертикальной химической стратификации глубоководных частей Каспия (табл. 4). В 1934 году были произведены весьма обширные сезонные океанографические работы на всем Каспийском море в ряде экспедиций. Из них особое значение имели зимние работы Академии Наук СССР (6 разрезов на Южном и Среднем Каспии, февраль-март 1934 г.), гидрохимическая часть которых проводилась Институтом рыбного хозяйства и океанографии (ВНИРО), и летняя океанографическая съемка всего Каспийского моря, произведенная ВНИРО на пяти судах единовременно под руководством С.В, Бруевича (август-октябрь 1934 г. - 29 разрезов по всему морю, 235 станций, 5900 миль пути).
В 1935 и 1936 гг. под руководством Бруевича работала Южно-Каспийская гидрохимическая экспедиция ВНИРО, чья задача заключалась в более детальном развитии отдельных проблем химии Каспийского моря.
Гидрологический облик Каспия
Морфологически Каспийское море обычно делится на три части: очень мелководный Северный Каспий и глубоководные Средний и Южный Каспий, разделяемые подводным Апшеронским порогом, над которым максимальная глубина составляет около 200 м. Средняя глубина Северного Каспия 6,2 м, максимальная глубина Среднего Каспия около 770 м и Южного несколько менее 1000 м. Границей между Северным и Средним Каспием принимается линия о. Чечень - мыс Тюб-Караган (п-ов Мангышлак), между Средним и Южным - линия о. Жилой (близ Апшеронского полуострова) - мыс Куулы (около 40°18 сш). Согласно проф. Н.М. Книповичу объемы и площади отдельных частей Каспия характеризуются следующими цифрами:

Таблица 1
Водный баланс Каспийского моря по Г.Р. Брегману и А.И. Михалевскому выражается следующим образом:
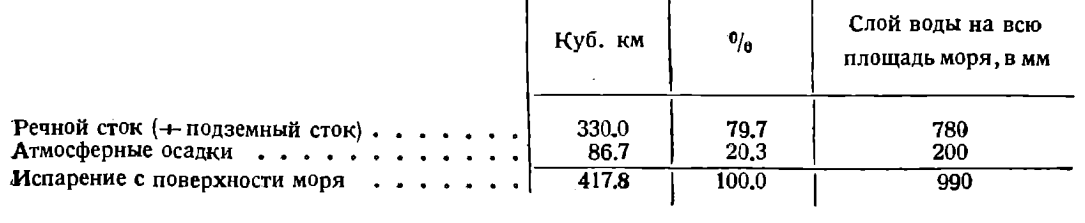
Таблица 2
Из всего речного стока в Каспий сток Волги составляет по различным данным от 76,3 до 81,4% от суммарного речного стока.
Течения в Каспийском море носят, согласно Н.М. Книповичу, круговой характер и направлены против часовой стрелки. Средний и Южный Каспий имеют свои циклы вращения, входящие в общий цикл движения воды во всем Каспии. Уровень воды Каспийского моря испытывает непрерывные колебания речного стока. Начиная с 1932 года Каспийское море претерпевает очень сильное продолжающееся падение уровня, не имевшее прецедентов за предыдущие 100 лет.
Высоты уровня моря, выраженные в сантиметрах над нулем Бакинского футштока (28,83 м ниже уровня Черного моря), за период 1830-1930 гг. имели максимум в 1869 г. - 387 см, минимум в 1925 г. - 263 см. После этого падение уровня представляется в следующих цифрах (в см): в 1930 г. уровень был 315, в 1931 г. - 303, в 1932 - 311, в 1933 - 308, в 1934 г. - 286, в 1935 г. - 267, в 1936 г. - 246, в 1937 г. 224 см. Средний уровень моря за 1830-1931 гг. - 326 см.
Осолонение верхних слоев Каспия, происходящее при падении уровня, вызывает увеличение вертикальной циркуляции моря, увеличение содержания биогенных элементов в верхнем слое моря и усиление биологической продукции моря.
Температура, соленость и пути аэрации моря
Распределение температуры на поверхности наиболее характерно зимой, когда меридиональное расположение моря обусловливает наибольшие разницы между северной и южной частями моря. Северный Каспий в это время в большей своей части покрыт льдом. Даже к югу температура от +1,0, +3,6° на границе Северного и Среднего Каспия повышается до +5° в северной части Среднего Каспия и далее на юг, особенно вдоль восточного мелководья. Максимальные температуры наблюдаются зимой в южной части Южного Каспия, где температура несколько выше +10°.
Область распространения повышенных зимних температур по всему Среднему и Южному Каспию весьма точно совпадает с областью распространения повышенных соленостей, обусловленных предшествующим усиленным летним испарением. И обратно низкие температуры связаны с пониженными соленостями.
Летнее распределение температур на всей поверхности Каспия довольно однородно и колеблется преимущественно в узких границах 23-26°, за исключением прибрежных мелководий, где температура может подниматься до 30-31°. Приходящие иногда с севера штормы понижают температуру на несколько градусов.
Вертикальное распределение температуры (табл. 3 и 4) указывает на большую однородность температур ниже 200 м. Температура воды в Среднем Каспии от 400 м и до дна (несколько менее 800 м) 4,82-4,89°, в Южном Каспии от 300 м и до дна - -5,90-5,96°. Сезонные колебания температуры ясно проявляются в верхнем 50-метровом слое и весьма незначительно - глубже. С температурными условиями непосредственно связаны вертикальная циркуляция и механизм аэрации глубинных вод Каспия. Нижняя граница вертикальной циркуляции, обусловленной зимним охлаждением, лежит в большинстве случаев между 50 и 100 м ниже поверхности, но в ряде случаев захватывает слой и мощнее 100 м. Однако область перемешивания распространяется несколько глубже области прямого влияния вертикальной циркуляции вследствие эффекта турбулентного перемешивания между слоем вертикальной циркуляции и нижележащими массами воды. Эта граница, изменяющаяся во времени и пространстве, лежит в среднем около 100 м глубины. Аэрация более глубоколежащих масс воды в Среднем Каспии происходит за счет энергичной вертикальной циркуляции в сильно охлаждаемой зимой северной части Среднего Каспия и сползанием охлажденных и аэрированных вод по северному склону Среднего Каспия в область больших глубин. В Южном Каспии дополнительная аэрация и охлаждение глубинных вод зависят от переливания более холодных и более аэрированных вод Среднего Каспия через Апшеронский порог. Средняя глубина распространения этих вод около 500 м, опускаясь иногда и значительно ниже. Другим фактором аэрации глубинных вод является зимнее сползание охлажденных и аэрированных вод высокой солености с восточных мелководий (Н.М. Книпович). Наконец, значительные на Каспий вертикальные перемещения водных масс под действием ветров также являются фактором периодического транспорта кислорода ко дну.
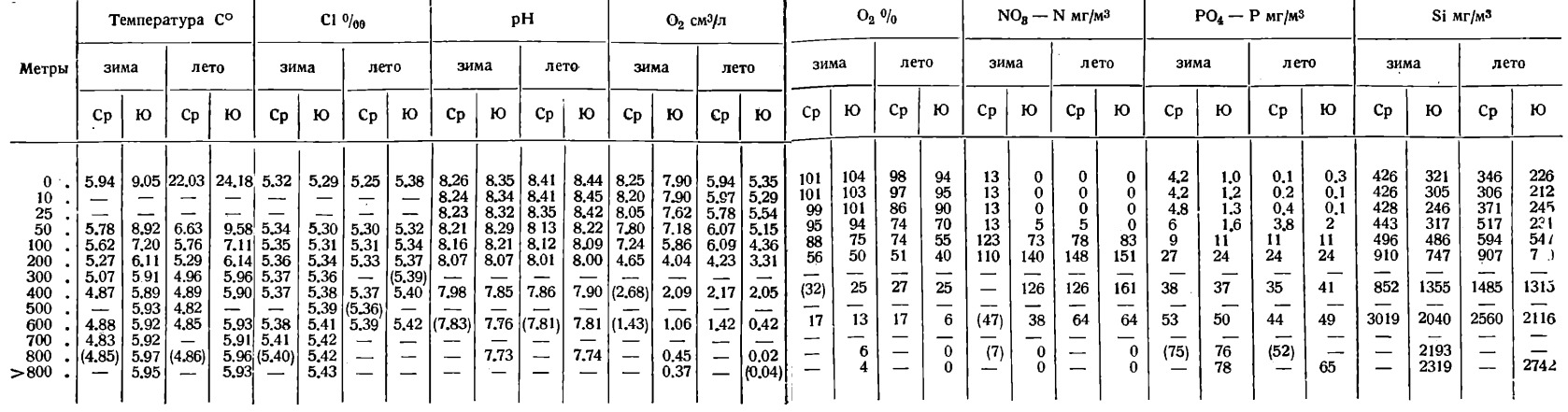
Таблица 3. Средние зимние и летние величины гидрохимических данных в Среднем (Ср) и Южном (Ю) Каспии, 1934 г.
Средняя величина хлора, в г/кг, на Среднем Каспии равна по нашему расчету 5,36‰, на Южном Каспий 5,38‰ и для всего моря в целом без Карабугаза – 5,37‰.
Это отвечает соответственным величинам солености 12,76, 12,80 и 12,78‰.
Сезонное распределение хлора на поверхности по данным 1934 г. было весьма своеобразным (табл. 3): зимой выше в Среднем Каспии, чем в Южном, летом - обратно. Сильно повышенные величины хлора приходятся зимой на восточное мелководье Среднего и Южного Каспия (Cl>5,35‰).
Пониженные величины хлора зимой отмечаются в северо-западной части и по западному берегу Среднего Каспия. Летом большие величины хлора обнаружены в западной и меньшие - в восточной части Среднего и Южного Каспия. В основном, по-видимому, эти сезонные различия в распределении хлора на поверхности связаны с темпами продвижения опресненных материковым стоком вод вдоль западного берега по схеме течений Н.М. Книповича. Вертикальное распределение хлора характеризуется очень малыми градиентами хлора по глубине. Обычно разница между содержанием хлора на поверхности в открытом море и на максимальной глубине не превышает 0,14‰ Cl.
Химическая стратификация
Химическая стратификация Каспия определяется динамикой водных масс и обменом между отдельными слоями. Основным фактором вертикального расчленения водных масс Каспия является температура, определяющая различия в плотности воды по вертикали. Влияние солености в этом отношении незначительно. Зимняя вертикальная циркуляция плюс турбулентное перемешивание на границе с нижележащими слоями воды обусловливает возможность перемешивания и хорошей аэрации лишь в верхнем 100-метровом слое. Слой этот является сильно отчлененным от всей нижней массы воды и характеризуется весьма низким содержанием биогенных элементов («питательных солей») и сравнительно высоким содержанием кислорода. Противоположными свойствами - ясно выраженным обогащением биогенными элементами и прогрессирующим падением кислорода обладает вся нижележащая масса воды. Первая область была названа зоной обеднения биогенными элементами, вторая - зоной аккумуляции биогенных элементов (табл. 4).
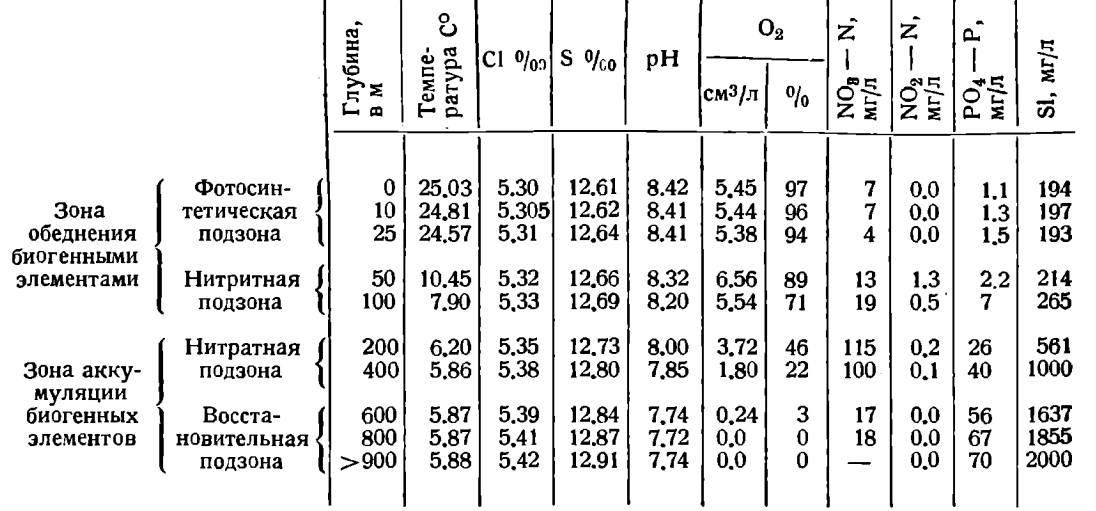
Таблица 4. Разрез Куринский Камень – о. Огурчинский (35°55 сев. шир.). 14-21 августа 1933 г.
Причиной обеднения верхнего слоя биогенными элементами является потеря их с падающими в нижнюю зону планктонными (преимущественно животными) организмами. Обратное поступление биогенных элементов из нижней зоны в верхнюю, так же как и поступление кислорода из верхней зоны в нижнюю, весьма затруднено и может происходить лишь в меру развития турбулентных процессов на границе этих двух зон. Постоянная динамическая устойчивость концентрации биогенных элементов в верхней зоне обусловливается только эффективностью турбулентного перемешивания на границе этих двух зон.
В пределах зоны обеднения естественно выделяется отдельная резко выраженная подзона фотосинтеза, нижняя граница которой располагается на Каспийском море преимущественно между 25 и 50 м и реже между 10 и 25 м. Слабые фотосинтетические процессы проявляются и глубже - на 50 м, но не имеют здесь определяющего значения. Потребление биогенных элементов в разгар вегетативного периода происходит главным образом в пределах подзоны фотосинтеза и лишь частично за счет того количества биогенных элементов, которое при летней стратификации плотностей может быть передано при турбулентном перемешивании из нижней части зоны обеднения. Зимнее перемешивание влечет новое обогащение верхнего слоя биогенными элементами в пределах его содержания во всей зоне обеднения и таким образом обусловливает постепенное вовлечение всех запасов биогенных элементов зоны обеднения в область фотосинтеза.
Процесс этот, принципиально одинаковый для Среднего и Южного Каспия, протекает, однако, в обеих частях Каспийского моря с различной силой. Зимнее, накопление, регенерация биогенных элементов, выраженное на Среднем Каспии довольно значительно, на Южном Каспии весьма мало. В этом отношении Южный Каспий близок к тропическим районам Мирового океана.
В нижней подзоне зоны обеднения, на глубинах 50-100 м, особенно на горизонте 50 м, наблюдается повышение содержания нитритов, сопряженное с процессами распада и минерализации здесь органического вещества отмерших планктонных организмов, падающих из фотосинтетического слоя. Это наличие нитритов в слое 50-100 м и дало повод назвать нижнюю область зоны обеднения нитритной подзоной.
В зоне аккумуляции с увеличением глубины наблюдается увеличение биогенных элементов и прогрессирующее уменьшение содержания кислорода с глубиной. Нитрификация аммиачного азота между 400-600 м уже резко задерживается, и минерализация органического азота задерживается на стадии аммонификации. Это обусловливает весьма характерное для Каспийского моря поясное распределение нитратов. Нитратная подзона зоны аккумуляции имеет верхнюю границу обычно выше 100 м и нижнюю приблизительно около 500 м. Нижняя подзона зоны аккумуляции, характерная резким уменьшением содержания кислорода до полного его отсутствия, была названа нами восстановительной подзоной. Граница между нитратной и восстановительной подзоной хорошо совпадает с пределом влияния сползающих с северного склона Среднего Каспия вод, аэрированных при зимней циркуляции.
В Южном Каспии эта граница совпадает со средним пределом влияния относительно лучше аэрированных вод Среднего Каспия, переливающихся через подводный Апшеронский порог и Южный Каспий.
Говоря о границе между зоной обеднения и зоной аккумуляции, следует указать, что эта граница была установлена по распределению фосфатов.
Граница повышения и концентрации нитратов лежит несколько выше: обычно на горизонте 100 м нитраты уже резко выражены. По большей части еще несколько выше расположена граница накопления кремнекислоты. Наиболее высоко расположена граница скачкообразного возрастания содержания углекислоты - непосредственно под фотосинтетическим слоем. Эта последовательность в границах зон накопления углерода, кремния, азота и фосфора точно отвечает убыванию содержания этих элементов в диатомовом планктоне и обусловлена, очевидно, мощностью отдачи данного биогенного элемента в воду при разрушении отмершего планктона.
Положение поверхности, разделяющей зону обеднения от зоны аккумуляции, несколько изменяется в различные сезоны. Зимой пограничная поверхность почти на всех разрезах имела куполообразный характер, повышаясь в центре и опускаясь по краям. Летом куполообразный характер пограничной поверхности наблюдался только как исключение. С другой стороны, летом наблюдается поднятие холодных глубинных вод к восточному берегу, не наблюдавшееся зимой. Это поднятие глубинных вод к восточному берегу, как результат северных ветров, дующих вдоль восточного берега, находится в хорошем согласии с теорией сгонов и нагонов Экмана.
Солевой состав и определение возраста современного Каспия
По солевому составу вода Каспийского моря представляет сильно метаморфизированную под действием речного стока воду океанского происхождения. Степень метаморфизации может быть характеризована величиной хлорного коэффициента - отношение суммы солей к хлору, равного, по данным А.А. Лебединцева, 2,38 против 1,809 для океанской воды с соленостью, равной каспийской. Наиболее достоверным из прежних анализов воды Каспийского моря являются 4 анализа А.А. Лебединцева и его учеников, среднее из которых приведено нами в табл. 5. В виду недостаточности этих данных, в 1933 г. были произведены в различных пунктах и на разных глубинах выемки 11 проб воды Южного Каспия, переданные для анализа О.П. Опариной и М.Т. Голубевой. Среднее из этих данных, представляющее наиболее точный состав воды Каспийского моря, дано в той же таблице 5. Наиболее точная величина хлорного коэффициента, определенная из этих данных, составляет Ѕ/СІ = 2,40. Эту величину рекомендуется употреблять лишь для точного подсчета количества солей, оставляя для сравнимости с прежним материалом для вычисления «гидрологической» солености коэффициент 2,38. Анализы Опариной и Голубевой указывают на очень большую однородность в солевом составе воды Южного Каспия в различных точках и практическое тождество воды на 0 м и на глубине 970 м. Ясно заметным оказалось лишь уменьшение содержания кальция и карбонатов на очень мелководных станциях восточного берега и особенно в Красноводском заливе. Причина лежит в выпадении углекислого кальция из сильно нагретой воды при воздействии твердых карбонатов донных отложений.
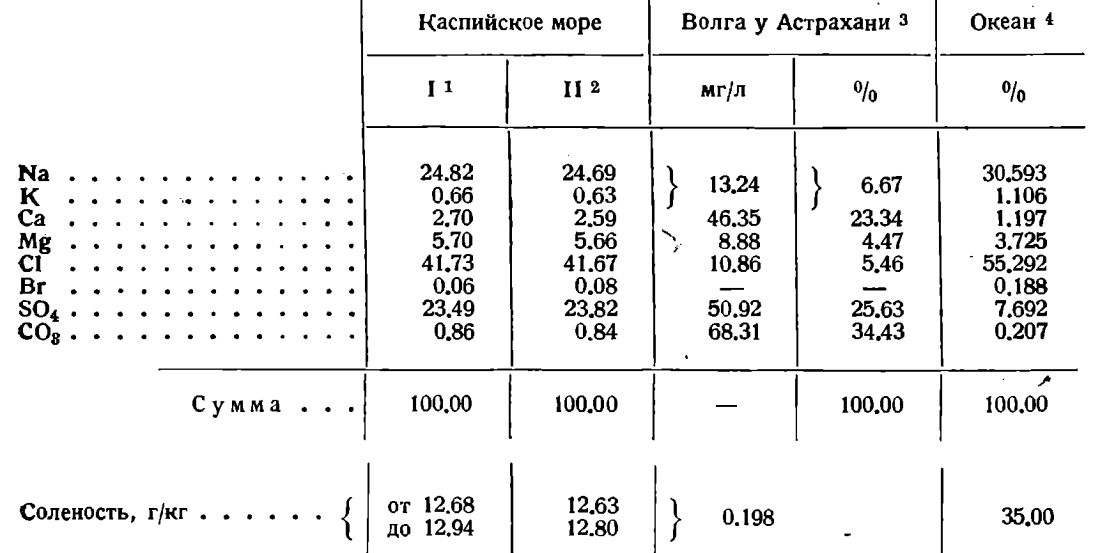
Таблица 5. Средний химический состав воды Каспийского моря и р. Волги.
При достаточном знании солевого состава воды Каспийского моря и воды впадающих в него рек возможно провести определение «возраста» современного Каспия, т.е. времени, понадобившегося для метаморфизации океанского солевого состава солями речного стока. Для Каспийского моря это определение «возраста» моря практически обозначает определение времени, прошедшего с последнего соединения древнего Каспия с Азово-Черноморским бассейном. Расчет производился следующим образом. Если обозначим через a и b общее содержание каких-либо двух ионов в море и через a' и b' валовое содержание этих же ионов в среднем годовом речном стоке, через n «возраст» моря в годах и через К отношение концентраций этих ионов в океанской воде, то из соотношения:
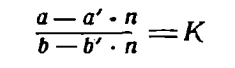
можем определить n. В качестве одного из ионов мы брали обычно хлор, в качестве второго сумму калия и натрия, магний или сульфаты. Вследствие отсутствия данных по анализам для всех рек, средний истинный состав р. Волги, подсчитанный по среднемесячным дебетам воды и среднемесячным анализам за каждый месяц, был принят как средний состав всего речного стока. Возраст современного Каспия, определенный таким образом, оказался равным по отношению CI/Na + K 6400 лет, по Сl/Ѕ04 - 10 600 лет и по Сl/Mg - 10 600 лет. Значительное расхождение между определениями по сумме щелочей и по магнию и сульфатам, по-видимому, связано с весьма ненадежным определением щелочей в речной воде. В качестве наиболее вероятного результата принимаем цифру 10 600 лет.
Последнее соединение Каспийского моря с Азовско-Черноморским бассейном относится, по предположению проф. П.А. Православлева, к началу голоцена. Последняя дата большинством геологов относится к времени от 4700 до 13 000 лет от наших дней. Таким образом, совпадение между геологическим и гидрохимическим расчетом возраста современного Каспия можно считать достаточно удовлетворительным.
Знание приближенного возраста Каспийского моря и баланса солей в нем позволяет подсчитать количество углекислого кальция, выделенного за время отчленения Каспийского моря от Азовско-Черноморского бассейна. Оно равно приблизительно одной тонне карбоната кальция на каждый квадратный метр площади моря: речной сток обусловливает отложение 94 г карбоната кальция на 1 кв. м ежегодно.
Ежегодное испарение воды в Карабугазском заливе обусловливает уменьшение солей, эквивалентное содержанию их в речном стоке за 3.7 года. Если принять во внимание выпадение карбоната кальция, то последняя цифра повышается до 8.6 лет.
Сток воды Каспийского моря в Карабугаз обусловливает ежегодное уменьшение солености моря на 0,0024‰ (S‰). Таким образом за время существования стока в Карабугаз повышение солевого состава моря временно прекратилось, сменившись уменьшением его.
Углекислота
Прямого определения давления углекислоты в воде Каспийского моря не производилось. Расчет по таблицам Буха и др. дал среднюю величину рСО2 около 4*10-4 атм., близкую для зимы и для лета. Лишь в течение исключительно интенсивного цветения диатомового планктона (Rhizosolenia calcaravis) в юго-восточной и восточной частях Южного Каспия в декабре 1935 г. вычисленное давление СО2 упало до 2,05 * 10-4 атм. Таким образом, если базироваться на расчете, то поверхность Каспийского моря в течение большей части года отдает угольную кислоту в атмосферу.
Эта отдача углекислоты в атмосферу может быть связана с громадным количеством углекислоты, освобождающимся в море при переходе гидрокарбонатов кальция речного стока в карбонаты с переходом последних в донные отложения. Количество это составляет для всего Каспийского моря около 1,8 * 107 т СО2. Принимая во внимание еще углерод органических веществ речного стока, а с другой - переход части органического вещества в донные отложения, можем считать, что море должно выделить в атмосферу ежегодно около 2 * 107 т углекислоты.
Это составляет приблизительно 2,3 куб. см на 1 кв. см в год. Количество это находится в пределах, допускаемых коэффициентом эвазии Бора, хотя следует заметить, что, по-видимому, вообще возможен обмен углекислоты между морем и атмосферой и в количествах, превышающих то, которое отвечает коэффициентам эвазии и инвазии Бора. Эта отдача морем в атмосферу половины количества углекислоты, поглощенной из атмосферы в процессах выветривания силикатный пород, представляет геохимический фактор универсального значения. Явление это должно быть более ярко выражено в морях, сильно подвергающихся влиянию речного стока. Превышение давления углекислоты в воде над атмосферным давлением СО2 в морях Каспийском, Черном и Балтийском даже в летнее время достаточно хорошо, подтверждает эту мысль.
Выпадение углекислого кальция из нагретых пересыщенных углекислым кальцием вод восточного мелководья было уже отмечено выше. Химически обнаруживаемое выпадение карбоната кальция находится в хорошем соответствии с исключительно высоким содержанием карбоната кальция в осадках этого района. Согласно опытам со встряхиванием воды с мелом все воды Южного Каспия от поверхности до дна (при глубинах почти до 1000 м) оказались пересыщенными карбонатом кальция - пресыщение это максимально для поверхностных вод мелководной зоны и минимально для придонных вод больших глубин.
Средняя величина щелочност и во всей толще воды Южного Каспия около 3,6 мг - экв./л; щелочной коэффициент А/СІ - около 0,675. Даже в области наибольших глубин щелочность от поверхности ко дну увеличивается не более как на 0,05-0,06 мг.- экв./л. Щелочность и щелочной коэффициент Каспийского моря выше, чем в океане и других морях, в том числе и в Черном море вследствие весьма энергичного воздействия речного стока.
Прочие определения
рН поверхностей воды Каспия в летнее время (табл. 1, 2) выше, чем рН океана и других морей, вследствие высокого щелочного резерва каспийской воды. Декабрьские наблюдения 1935 г. показали, что во время массового цветения Rhizosolenia pH поверхностных вод открытой части моря доходит до 8,60 - это максимально найденные величины для открытой глубоководной части Каспия.
Распределение кислорода в поверхностном слое зимой обнаруживает сильное убывание с севера к югу, что связано с понижением температуры на севере и повышением ее на юге в связи с резко выраженным меридиональным простиранием моря. Пути проникания в воду кислорода были указаны выше. Вертикальное распределение см. табл. 3 и 4. В кислородном обмене Каспийского моря весьма интересны максимумы кислорода ниже поверхности. Максимумы в Южном Каспии наблюдались на горизонтах 25 и 50 м, в Среднем Каспии обычно на горизонтах 50-100 м. Максимумы эти представляют реликты обогащения кислорода при зимней вертикальной циркуляции и вполне аналогичны таким же максимумам кислорода в арктических водах, в «холодном промежуточном слое».
Сероводород определялся нами только в Южном Каспий, где он находился лишь в области больших глубин, от 600-700 м и ниже. Содержание сероводорода в придонных слоях северной впадины Южного Каспия около 0,2 куб. см Н2Ѕ/л, в южной впадине до 0,29 куб. м. По данным Жузе содержание сероводорода в глубинных водах Среднего Каспия доходит до 0,51 куб. см. По данным А.А. Лебединцева (экспедиция 1904 г.), содержание сероводорода в глубинных водах Среднего Каспия доходит до 0,40 куб. см. Таким образом согласно этим авторам содержание сероводорода в глубинных водах Среднего Каспия заметно выше, чем в Южном Каспии. Сопоставление количества сероводорода с количеством альбуминоидного азота указывает на то, что лишь около 1/5 наличного количества сероводорода может быть отнесено за счет серы органических веществ, разлагающихся в анаэробных условиях. Остальная часть должна быть отнесена за счет восстановления анаэробного сульфатов.
Содержание аммиачного азота в августе 1934 г. определялось лишь на границе Северного и Среднего Каспия и было найдено максимальным у западного берега, в области влияния волжского стока, и у берега Мангышлака 39-40 мг N/м3 и минимальным в средней открытой части моря - 17-21 мг N/м3. В августе 1933 г. в Южном Каспии величины аммиачного азота колебались в различных пунктах от 20 до 130 мг N/м3, преимущественно же в пределах 40-70 мг/м3.
Нитриты зимой обнаруживались в Среднем Каспии в незначительном количестве в слое от 0 до 50-100 м, ниже нитриты отсутствовали (табл. 4). В Южном Каспии нитриты обнаруживали заметное накопление на горизонте около 50 м (0,1-0,5 мг N/м3 и до 1,0 мг N/м3) и отчасти 100 м и убывают и далее исчезают книзу.
Летом 1933 г. в Южном Каспии нитриты показали ясное накопление в количестве в среднем 1,3 мг N/м3 на глубине 50 м (табл. 4). То же наблюдалось летом 1934 г. в Среднем Каспии. Обе картины - зимняя и летняя являются типичными для моря.
Нитраты в Каспийском море, как указывалось выше, имеют весьма характерное поясное распределение и содержатся в слое от 100 до со 500 м в количестве 100-150 мг N/м3. Выше 100 м нитраты отсутствуют или находятся в незначительных количествах вследствие вовлечения их в зону фотосинтеза, где они потребляются, ниже 500 м вследствие восстановления в аммиак (Табл. 3).
Распределение фосфора и кремния – в таблице 3 и 4.
Весьма характерна картина зимнего распределения кремния в поверхностном слое, ясно указывающая на области распределения речного стока. Содержание кремния от 500-550 мг Р/м3 у западного берега планомерно понижается к восточному берегу до 200-300 мг Р/м3 в среднем Каспии и до 100 мг Р/м3 в Южном.
Суточные колебания кислорода (через 3 часа) определялись главным образом с целью дальнейшего вычисления суточной продукции фитопланктона. Суточные колебания кислорода в поверхностном слое летом значительно уступают суточным колебаниям в чистых реках средней полосы СССР и приблизительно таковы же, как на других морях. На глубине 25 м суточные колебания летом достигают весьма больших величин приблизительно того же порядка, как и суточные колебания в незагрязненных реках средней полосы СССР. На поверхности суточные колебания кислорода ∆O2 max в Южном Каспии были равны летом 1933 г. 0,34 куб. см, в открытой части Северного Каспия - от 0,38-0,52 куб. см летом до 0,11 куб. см в октябре 1934 г. На глубине 25 м в Южном Каспии ∆O2 max было летом 1933 и 1934 гг. 0,85-1,12 куб. см. В заливе Мертвый Култук. в августе 1934 г. – 1,54 куб. см.
При определении продукции фитопланктона по гидрохимическим данным мы под продукцией фитопланктона подразумевали не прирост биомассы за данное время, но сумму генераций фитопланктона, образовавшихся за данное время в значительной степени из одних и тех же химических элементов. В качестве элементарного отрезка времени, служащего для химического определения продукции, принимаются 1 сутки. Вследствие обычного нарушения однородности воды в течение суток (течения) применение Пюттеровского принципа расчета может повести к значительным ошибкам в расчете продукции фитопланктона, и мы вели расчет по следующей формуле:
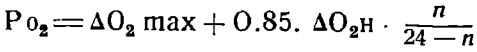
, где РО2 - полная фотосинтетическая продукция кислорода за день в см3/л, ∆O2 max - разница между послеполуденным максимальным содержанием кислорода и ночным (около восхода солнца) минимумом его в воде, ∆O2Н - убыль кислорода в воде в течение ночи, - от захода до восхода, n - продолжительность дня в часах. В тех случаях, когда вследствие неоднородности воды в море ошибки в определении ∆O2Н были чрезмерно велики, PO2 определялось по эмпирической формуле: РО2 = ∆O2 max * 1,6. Пересчет в глюкозу производился согласно соотношениям: 1 мг О2 = 0,94 мг глюкозы; 1 куб. см О2 = 1.34 мг глюкозы. Продукция кислорода в 1933 и 1934 гг. определялась на 9 суточных станциях и дала следующие результаты. Продукция фитопланктона, выраженная в миллиграммах глюкозы на литр воды, была равна - в открытой части Южного Каспия летом 1933 г. на поверхности 0,75, на 25 м – 1,82 – 2,50 (1933 и 1934 гг.). Зимой 1934 г. на поверхности 0,76, на глубине 25 м – 0,30; в Северном Каспии на поверхности в августе 1934 г. – 0,74 – 1,42, в сентябре-октябре – 0,56-0,19.
Отношение убыли отдельных биогенных элементов от зимы к лету в Среднем Каспии, где сезонная убыль представляет достаточно ощутимую величину, с учетом солевых поправок равно:
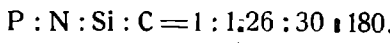
Для Ламанша это же соотношение было подсчитано нами из данных Аткинса и Купера по той же методике и найдено равным приблизительно 1:7,4:11,6:92. Приблизительное соотношение этих элементов в диатомовом планктоне равно 1:5,6:24,2:30,6; то же для перидиниевого – 1:8,2:5,4:60. Высокая трата углерода в Каспийском море по сравнению с содержанием его в фитопланктоне, возможно, связана с отдачей СО2 поверхностью моря в атмосферу.
Принимая среднее содержание фосфора в диатомовом планктоне в 2,38% Р2О5 и кремния в 31,5% SiO2 и содержание сухого вещества в сыром планктоне в 10%, получаем условный прирост биомассы фитопланктона в Среднем Каспии от зимы к лету равным в среднем 2 г/м3 сухого и 20 г/м3 сырого вещества («урожай», «crop» предыдущих исследователей).
Чрезвычайно характерна для биологической динамики данного водоема величина, названная «оборачиваемостью» фитопланктона: отношение продукции фитопланктона к его биомассе (Р/В - коэффициент проф. Л.А. Зенкевича). Для различных частей Каспийского моря величина эта была нами определена равной 2,8-3,7. Слабым местом в этом определении являлось неточное знание биомассы фитопланктона.
